
1. 서론
쓰레기 매립은 쓰레기의 소각이나 퇴비처리에 대한 대안으로 사용되는 처리방법이다. 쓰레기매립지는 침출수에 의한 수질오염, 매립가스, 토양침하 등으로 인해 생태적 교란이 발생하는 곳이며, 토양 특성에 따라 식생 변화가 생길 수 있다(Kang et al., 2015). 서울시 청소사업본부는 대규모 쓰레기매립지를 물색 중 서울시 외곽지이면서 교통이 편리하고 제방축조 공사 중인 난지도를 대상지로 선정하였다. 난지도 매립지는 1978년 3월부터 1993년까지 15년 동안의 쓰레기 매립이 이루어졌다. 제1매립지는 5,640만m3, 제2매립지는 3,480m3, 슬러지 매립지는 80만m3 등 총 9,200만m3의 쓰레기가 매립되었다. 이후 이 지역은 1996년부터 2002년까지 매립지 안정화 사업을 진행하였다(Seoul Metropolitan Government, 2003).
쓰레기매립지 식생은 쓰레기 출처에 따라서 기원이 다른 식물이 침입하여 분포하게 되고, 3년이 지난 매립지에서는 도심지에서 나타나는 식물의 피도가 커지며, 7-8년이 지나면 식생의 다양성이 줄어들고 인근 임야나 초지에서 나타나는 자연식생이 발달하게 된다(Maurice, 1998). 쓰레기 매립 후 출현하는 목본식물로 자작나무과의 Betula pubescens, 버드나무과의 Salix carprea(Ettala, 1988), 콩과의 Acacia confusa(Chan et al., 1997) 등이 있으며, 매립지에서 가장 많이 생육하는 식물은 콩과식물이다(Chan et al., 1998). 북미 애팔래치아 산맥에 분포 중심지를 가지고 있는 아까시나무는 북반구 온대 지역에 널리 식재된 자원 식물이다(Flint, 1997). 쓰레기매립지에서 아까시나무가 가장 넓은 면적을 차지하고 있는데, 이는 건조에 강하고, 질소고정능력이 있으며, 초기의 빠른 정착과 생육, 교란지역에서도 맹아 발달이 왕성하기 때문이다. 귀화식물 이외에도 환삼덩굴 등과 같은 덩굴성 식물들은 쓰레기매립지에서 피도가 크고 매립지 내 많은 지역에서 잘 자란다. 이들은 오염된 지역 근처에서 뿌리를 내리고, 오염된 지역은 접근하지 않으면서 위로 자라 다른 식물을 감아 올라가면서 빛 등의 환경요인에 대해 우위를 차지하기 때문이다(Wong and Yu, 1989).
우리나라 쓰레기매립지에 분포하는 식생군락의 분포 특성 및 변화에 관한 연구는 많지 않다. Lee et al.(1997)은 난지도 쓰레기 매립이 완료된 1993년 이후 3년이 지난 1996년에 매립지 사면 191ha 중 31ha가 목본식물로 천이가 진행하였다고 하였다. Kim(2008)은 난지도 쓰레기매립지 사면의 목본식생구조 분석 결과 아까시나무군락, 가죽나무-아까시나무-참오동군락, 능수버들군락으로 분류되었으며, 잠재자연식생이 전혀 출현하지 않아 생태적 천이 예측은 불가능하다고 하였고, 아까시나무와 능수버들은 선구식물로서의 역할을 하고 있음을 확인하였다. 최근 연구로 노을공원 쓰레기매립지에 이입된 아까시나무군락은 일반 도시림과 달리 참나무림으로 천이가 진행되지 못하고 조류 산포종인 뽕나무 등과 경쟁을 하는 것으로 분석되었다(Wei and Oh, 2021). 쓰레기에 의한 침출수, 매립가스, 복토층, 자연 침하 등 쓰레기매립지만의 독특한 식생기반은 자연상태와는 다른 난지도 쓰레기매립지만의 독특한 식생구조를 가지게 된다. 매립지 토양은 토양 내 산소공급, 양이온교환능력, 영양물질상태, 수분보유능력 등이 낮고, 높은 토양온도, 과다한 다짐 등으로 토양환경 또한 불량한 것으로 알려져 있다(Kim et al., 2001; Choi et al., 2002). 본 연구는 난지도 쓰레기매립지만의 독특한 식생구조를 밝히고자 23년간(1999-2021년) 현존식생과 식물군집구조의 현황 및 변화 특성을 분석하였다. 이 연구는 도시지역 쓰레기매립지 식생복원을 위한 기초자료 제공을 목적으로 수행하였다.
2. 연구방법
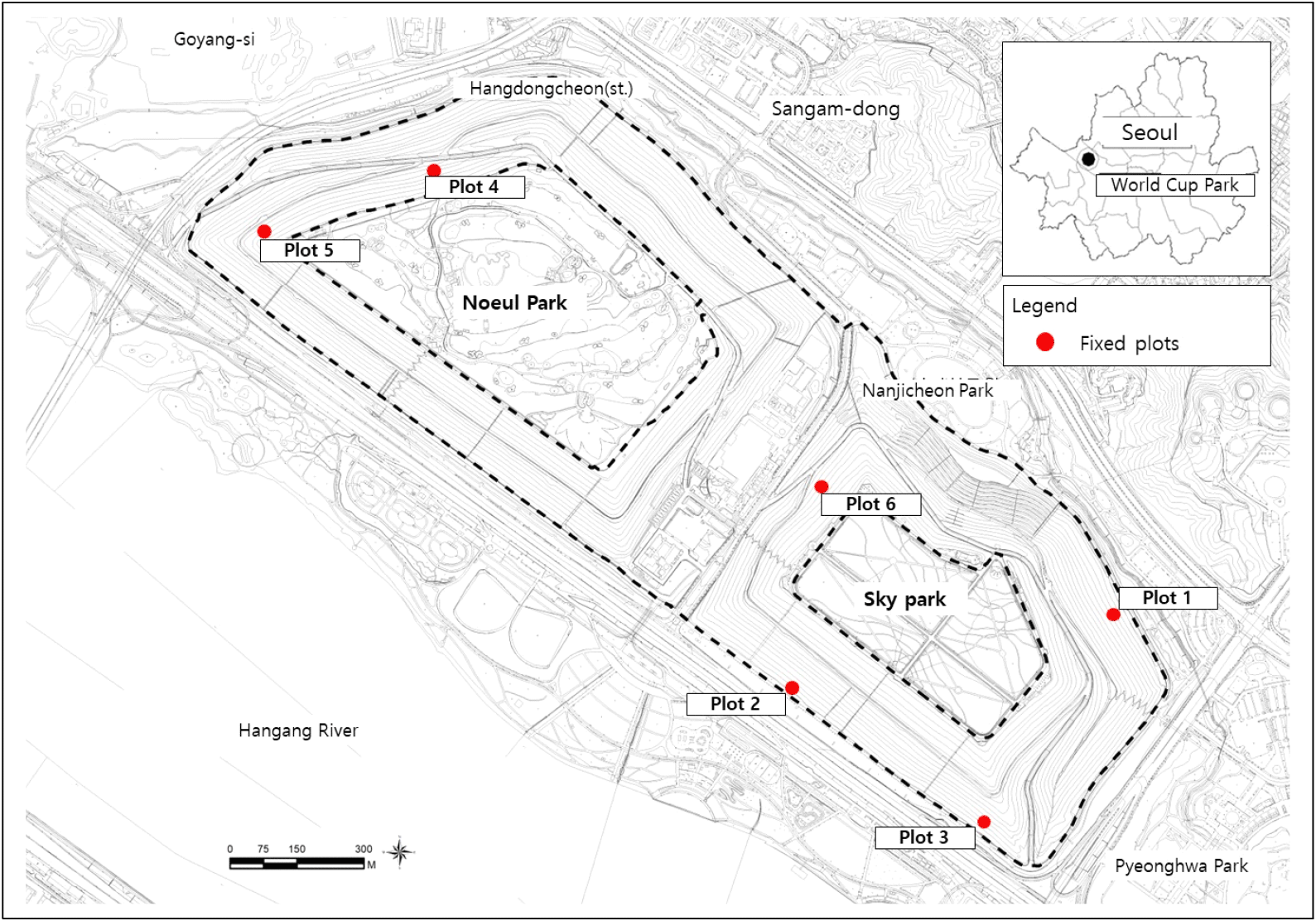
연구대상지는 서울시 마포구 상암동 478-1 일대에 위치하고 있다(Figure 1 참조). 1978년 3월부터 1993년 3월까지 쓰레기매립지였던 난지도는 2002년 월드컵공원으로 조성되었으며 하늘공원, 노을공원, 난지천공원, 평화의공원으로 나누어진다. 연구대상지는 월드컵공원 매립지 상부 노을공원과 하늘공원을 제외한 사면지구 1,126,029m2를 대상으로 현장조사를 실시하였다. 노을공원 사면과 하늘공원 사면은 탐방객의 출입을 통제하는 지역으로 인위적 간섭 없이 쓰레기 매립이 완료된 이후로 유지되고 있는 지역이다. 따라서 매립지 사면지역은 쓰레기매립지만의 독특한 식생구조가 발달해 있다고 할 수 있다. 특이사항으로 노을공원 시민모임은 최근 10년 동안 사면부에 소나무, 스트로브잣나무, 상수리나무, 꾸지나무 등을 식재하였다(Introduction to Friends of Noeul Park, 2020).
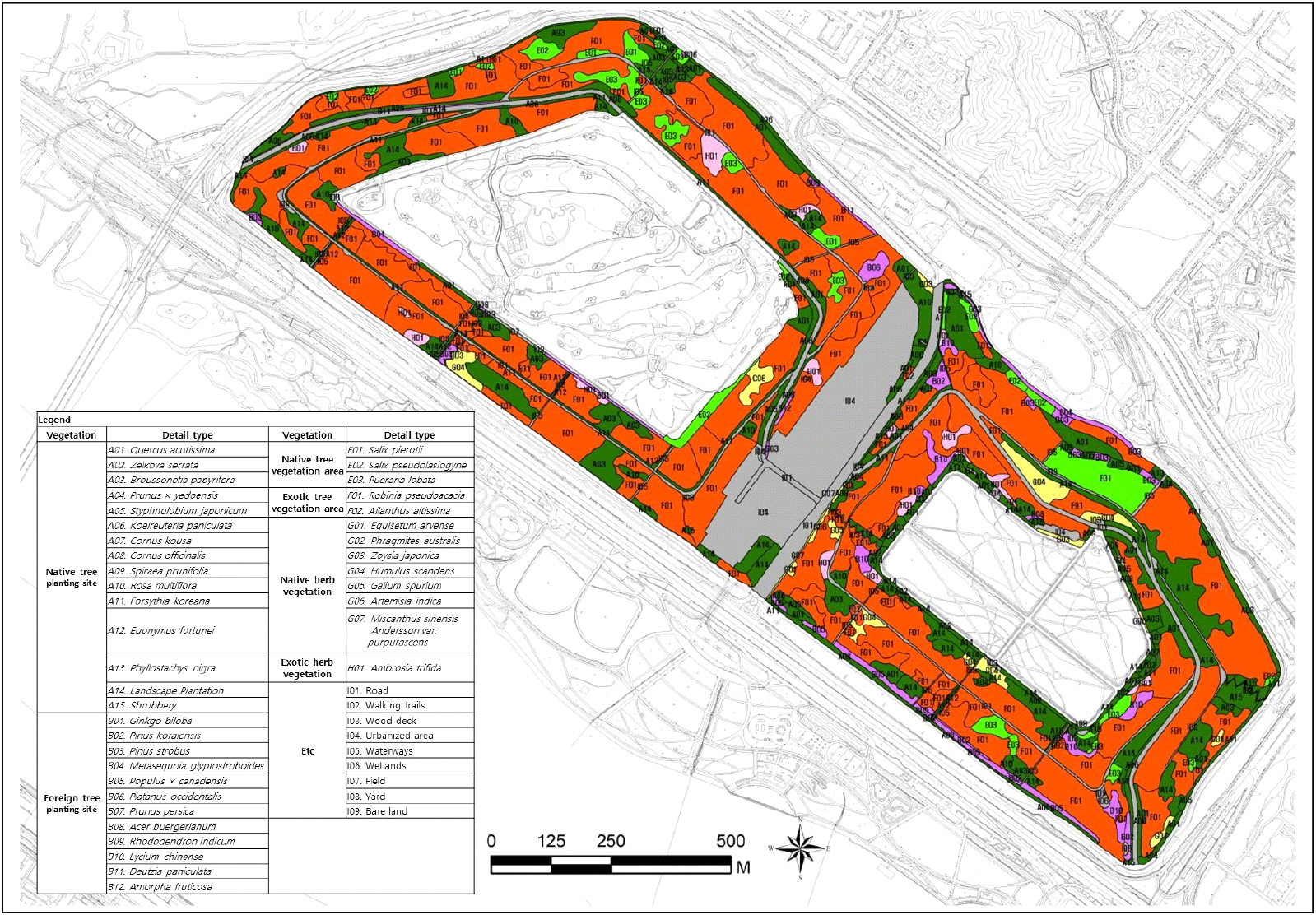
현존식생 조사는 사면지역의 식생상관(vegetational physiognomy)을 기본으로 현존하는 식물군락 유형별 분포현황을 1/1,000 수치지도를 이용하여 도면화하였다. 실내에서는 Auto CAD 2023과 Arc-map 10.8 프로그램을 활용하여 현존식생도 작성과 현존식생 유형별 면적 및 비율을 산출하였다. 현존식생 변화 분석은 1999년, 2003년, 2005년, 2007년, 2008년, 2012년, 2016년, 2021년 현장조사 결과를 비교하여 파악하였다. 현존식생 유형은 크게 식재종과 자연발생종으로 구분하였으며, 식재종은 매립이 완료된 1993년 이후로 매립지 사면지역에 인위적으로 식재한 종이고, 자연발생종은 매립이 완료되기 전부터 현재까지 자연발생적으로 정착하여 분포하는 종을 말한다. 식재종과 자연발생종은 매립지 사면의 독특한 생육기반조건에 따라 초본생육지역과 목본생육지역으로 분류하여 분석하였다. 그리고 자생 여부에 따라 자생종과 외래종으로 구분하였다. 자생식물은 외부에서 자연적으로 이입된 자생종뿐만 아니라 식재한 자생종도 포함하였으며, 외래식물은 귀화식물을 비롯하여 원예종과 재배종을 포함하였다.
식물군집구조 조사는 Monk et al.(1969)의 방법을 참고로 하여 흉고직경(DBH) 2cm 이상의 수목군을 교목층, 2cm 이하를 관목층으로 하였다. 현장조사는 수종명, 흉고직경(cm), 수고(m), 지하고(m), 수관폭(m × m)을 측정하여 기록하였다. 식물군집구조 조사는 자연발생림 내 1999년에 설정한 고정조사구 총 6개소를 현장조사하였다. 고정조사구는 대상지 내 넓게 분포하고 있는 아까시나무군락, 소규모 군락을 형성하고 있는 능수버들, 참오동나무-가죽나무를 대상으로 밀도와 경급을 고려하여 선정하였다. 고정조사구는 아까시나무군락 2개소, 가죽나무-아까시나무-참오동나무군락은 1개소, 능수버들군락은 3개소이다(Table 1 참조). 1999-2008년까지는 교목층 수고를 고려하여 각 조사구 면적을 100m2로 설정하였으나 2012년 이후 자연발생림에 설정된 고정조사구는 20m × 20m(400m2)를 기준으로 설정하였다. 따라서 연도별 변화상의 분석은 과거 조사구와의 면적 기준을 일치시키기 위하여 20m × 20m 조사구 중 과거 조사구에 해당하는 10m × 10m 조사구만을 분리하여 식물군집구조 변화를 분석하였다.
각 수종의 상대적 우세를 비교하기 위해 Curtis and McIntosh(1951)의 중요치(importance value; I.V.)를 통합하여 백분율로 나타낸 상대우점치(Brower and Zar, 1977)를 수관층위별로 분석하였다. 상대우점치(importance percentage; I.P.)는 (상대밀도 + 상대피도) / 2로 계산하였으며, 개체 간 크기를 고려해 수관층위별로 가중치를 부여한 (교목층 I.P. × 3+아교목층 I.P. × 2 + 관목층 I.P. × 1) / 6으로 평균상대우점치(mean importance percentage; M.I.P.)를 산정하였다(Park, 1985).
식생조사 자료를 바탕으로 조사구별 종수 및 개체수를 비교․분석하였다. 종다양도지수는 종풍부도와 종균등도를 하나의 수치로 나타낸 것이다. Shannon의 종다양도(H’)는 Shannon and Weaver(1949)가 제시한 것으로 군집 연구에 가장 많이 이용되고 있으며 희소종의 중요성을 인정해주는 지수로 아래와 같은 식을 이용하여 구하였다. 이 외에도 최대종다양도(H’max), 균제도(J’), 우점도(D)를 구하였다. 아래 수식의 ni는 전체 종의 총 개체수에 대한 어떤 종의 개체수 비이고, S는 구성종수를 나타낸다.
▪ H’ = −∑ (ni / N) log ni/N
▪ H’max = log S
▪ J’ = H’ / H’max
▪D = 1 - J’
3. 결과 및 고찰
월드컵 사면지구 현존식생을 분석한 결과 자생목본식재지 22.1%, 외래목본발생지 51.4%, 외래목본식재지 3.4%, 자생목본발생지 4.4%, 자생초본발생지 2.0%, 외래초본발생지 1.6%, 기타 15.3%로 분포하였다(Table 2, Figure 2 참조). 자생목본식재지는 상수리나무 3.3%, 꾸지나무 2.8%, 모감주나무 2.8%, 조경수식재지 7.6% 등이 주요 식재종이었고, 개나리, 찔레꽃, 왕벚나무, 산수유, 조팝나무 등이 함께 식재되어 있었다. 식재종은 대부분 주요 동선 주변, 접근성이 양호한 사면 하단부 등에 식재되어 있었다. 외래목본식재지는 구기자나무 0.9%, 이태리포플러 0.7%, 스트로브잣나무 0.4% 순이었고, 은행나무, 잣나무, 메타세콰이아 등이 식재되어 있었다. 외래목본발생지는 아까시나무가 51.4%로 대부분이었고, 가죽나무는 소규모 면적으로 분포하였다. 자생초본 발생지는 환삼덩굴 1.3%, 쑥 0.3%이었고, 잔디, 억새, 갈퀴덩굴 등이 함께 생육하고 있었다. 외래초본 발생지로 단풍잎돼지풀은 전체 면적의 1.6% 분포하였다.
월드컵공원 사면지구의 1999년부터 2021년까지 23년간 현존식생 변화를 살펴보면 목본식재지는 지속적으로 증가하였는데, 특히 자생목본식재지는 1999년 0.8%를 시작으로 지속적으로 증가하였다. 2008년 18.2%이었으나 2012년 10.7%로 감소 후 최근 노을공원 100개숲 만들기 사업을 통해 상수리나무, 꾸지나무, 왕벚나무, 모감주나무 등 식재면적이 증가하여 2021년 22.1%로 확인되었다. 외래목본식재지는 2003년 1.4%로 시작하여 2007년 4.0%로 증가하였으나 2012년 1.8%로 감소하였고, 2021년 꽃말발도리, 영산홍 등이 추가 식재되어 3.4%로 증가하였으나 전체 면적 변화는 미미하였다. 자연발생숲은 대부분 외래목본의 아까시나무림이었으며, 1999년 17.3%로 조사된 이후 2008년 35.4%로 1999년 대비 두 배 이상 증가하였으나 2012년 33.9%, 2016년 33.3%로 감소하다 2021년에는 가죽나무 관리 및 아까시나무 재발생에 따라 51.4%로 크게 증가하였다. 자생목본의 경우 1999년에는 외래목본과 비슷한 수준인 12.0%의 면적으로 군락을 형성하였는데 2008년 18.0%까지 다소 증가한 이후 2012년 6.3%로 크게감소하였으며, 2016년에는 10.9%로 증가하다 2021년 버드나무, 능수버들의 세력 감소에 따라 4.4%로 조사되었다. 자연발생초본의 경우 1999년에는 외래초본 1.6%, 자생초본 32.0%로 자생초본이 넓은 면적을 차지하였으나 이후 자생초본의 지속적 감소와 외래초본의 증가, 상층 교목 세력 증가로 인해 외래초본은 2012년 8.3%, 2016년 7.4%, 2021년 1.4%로 분포하였고, 자생초본은 2005년 16.4%, 2016년 5.7%, 2021년 2.0%로 분포면적이 감소하였다(Table 3, Figure 3 참조).
 Native tree planting
Native tree planting  Exotic trees naturally occurring
Exotic trees naturally occurring  Native tree naturally occurring
Native tree naturally occurring 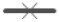 Native herb naturally occurring
Native herb naturally occurring
생육특성별 현존식생 면적을 분석한 결과, 초본식생에서 목본식생으로 천이가 진행 중에 있었으며 자연발생한 자생종은 감소하고 아까시나무를 비롯한 귀화식물의 면적이 증가하였다. 주요 변화상으로 2008-2012년을 기점으로 면적의 증감이 크게 발생하였는데, 이는 2010년 강풍을 동반하며 한반도를 통과한 태풍 곤파스(Kompasu)의 영향으로 판단되었다. 제7호 태풍 곤파스는 2010년 8월 29일 21시에 일본 오키나와 남동쪽 약 880km 부근 해상에서 강도는 약, 크기는 소형인 태풍으로 발생, 이후 해수면 온도가 28-29℃ 이상인 해역을 북서진하면서 8월 31일 10시에는 강도는 강, 크기는 중형인 태풍으로 발달하였다(Korea Meteorological Administration Press Release, 2010).
이 태풍은 비보다는 강한 바람으로 위력을 떨친 ‘마른 태풍’으로 9월 2일 6시 35분 강화도에 상륙하여 약 5시간여만에 우리나라 중부지역을 관통하여 서울, 경기도, 인천, 충청남도 및 전라남도의 총 89개소에서 20,107본의 가로수를 쓰러트렸으며, 총 37억 3,656만 원의 재산피해를 입혔다(Youn et al., 2011). 이 과정에서 노을공원 매립지 사면 내에서도 많은 풍도목이 발생하였다. 풍도목(風倒木) 또는 풍도(風倒)는 기후, 지형, 임분, 입목 및 토양 인자 등의 상호 작용에 기인하여 바람에 의해 뿌리가 뽑히거나 수간이 파괴된 입목을 뜻하며(Mitchell, 2001), 태풍 또는 강풍에 의해 발생하는 자연현상이다. 특히 사면지역은 건설폐자재의 매립과 다짐으로 형성된 단단한 지반 위에 복토를 통해 식재기반이 형성된 지역으로 토심이 얕고 토질이 불량한 지역이다. 사면지구의 수목은 강풍으로 인해 도복되면서 분포면적과 세력 변화가 발생하였고, 특히 건조하고 불량한 식재기반 위에서 군락을 형성하던 습지성 목본인 버드나무는 세력이 감소하게 되었다. 초본의 변화는 이러한 목본의 도복 및 쇠퇴와 밀접한 연관성이 있는데, 수목에 의해 울폐되어있던 지역이 개방되면서 단풍잎돼지풀이 급격하게 분포면적을 확대한 것으로 판단되었다. 자생초본 중 환삼덩굴은 단풍잎돼지풀과 최근 유입된 가시박 등이 확산되면서 분포면적이 감소하였다.
아까시나무는 건조에 강하고 생장이 빠르며 질소를 고정하는 능력이 있어(Converse and Betters, 1995) 생태적 지위가 넓은 종으로 알려져 있다. 그리고 공중질소고정으로 영양분을 보충할 수 있기 때문에 토양이 극도로 건조하고 영양분이 적은 곳에서도 생육이 가능하다. 교란된 지역에서 뿌리가 상당한 맹아를 만들어 내어 맹아력이 강하며(Boring and Swank, 1984; Hong and Song, 1990) 인접 임분의 종류에 관계없이 수평근의 맹아를 이용하여 전 방위로 이입하고 있어(Yun et al., 2001) 서울시 도시공원 내 산림 가운데 가장 넓은 면적을 차지하고 있다(Lee et al., 1993).
아까시나무군락 고정조사구 1의 층위별 상대우점치 분석결과 아까시나무가 교목층에서 100.0%, 아교목층에서 89.9%로 우점하였고, 아교목층에 아까시나무 이외 수종으로 벚나무가 10.2%로 출현하였다. 관목층에서는 찔레꽃의 우점도가 88.9%로 높았으며 아까시나무 7.7%, 산수유 3.4%로 함께 출현하였다. 출현 종수 및 개체수는 4종 94개체로 종수가 매우 적었으며, 우점종인 아까시나무의 우점도가 높아 샤논의 종다양도지수는 0.381로 매우 낮았다(Table 4 참조). 군집구조 분석결과를 종합해보면 교목층, 아교목층에서 아까시나무가 우점하고 일부 자연발생된 벚나무가 아교목층에서 출현하였으나 우점도가 매우 낮았다. 따라서 지속적으로 아까시나무림이 유지될 것으로 판단되었다.아까시나무군락 고정조사구 2의 층위별 상대우점치 분석결과 상수리나무가 교목층에서 100.0%로 우점하였고, 아교목층에 밤나무 42.2%, 팽나무 31.0%, 붉나무 26.8% 순으로 출현하였다. 관목층에서는 찔레꽃이 37.2%, 칡이 28.0%로 높은 우점도를 보인 가운데 뽕나무, 팽나무, 참느릅나무 등이 함께 출현하였다. 출현 종수 및 개체수는 13종 137개체로 샤논의 종다양도지수도 0.9684로 아까시나무군락에 설정된 다른 조사구에 비해 높은 수치를 보였다(Table 5 참조). 군집구조 분석결과를 종합해보면 교목층에서 상수리나무가 우점하고 아교목층에서 밤나무, 팽나무, 붉나무가 출현하였으며, 관목층에 다양한 종이 출현하였다. 아교목층과 관목층에 교목성상의 수종이 발생하여 추후 식생구조 변화가 예측되었다. 아까시나무군락 고정조사구를 대상으로 종합적인 식생 변화 경향을 분석한 결과 목본 종수는 2008년까지 5-7종을 유지하였으나 2012년 2종으로 크게 감소하였고, 2016년에는 4종, 2021년 11종으로 크게 증가하였다. 이러한 변화는 2010년 태풍에 의한 풍도목 발생, 아까시나무 세력 확장 등으로 인한 것으로 판단되며 이후 유입종들에 의해 종수는 지속적으로 변화할 것으로 판단되었다. 특히 교목층과 아교목층 개체수의 경우 2008년까지 교목층 7.5-10.0개체, 아교목층 7.6-12.9개체가 유지되었으나 2012년 교목층 3.9개체, 아교목층 4.9개체, 2021년 교목층 4.5개체, 아교목층 6.0개체로 2008-2012년 사이 교목층과 아교목층의 개체수가 크게 감소한 이후 2021년까지 다시 증가한 것으로 나타났다(Table 6 참조). 샤논의 종다양도지수는 2012년까지 큰 변화가 없었으나 2016년 0.3703, 2021년 0.9546으로 크게 증가하였다(Table 7 참조). 교목층과 아교목층 밀도 감소로 인해 상층이 개방되면서 다양한 유입종의 발생에 따라 지속적으로 증가한 것으로 판단되었다.
아까시나무군락의 천이 예측 관련 선행연구로 Yun et al.(1999)은 아까시나무 조림지에서 천이 예측과 조림학적 제어에 관한 연구에서 크게 5가지 군락으로 구분하면서, 아까시나무 조림지는 참나무류, 단풍나무류, 벚나무류 등이 혼재하는 산림으로 천이가 진행될 것이라고 예측하였다. 하지만 본 군집은 매립지 식재기반의 특수성과 함께 대기오염 물질의 지속적인 공급 영향 등으로 인해 Cho and Kim(2005)의 연구 결과처럼 아까시나무의 짧은 수명과 질소고정 박테리아와의 공생에 따른 천근성으로 노령 개체의 도복 현상, 태풍에 의한 도복, 그에 따른 숲 틈이 발생하지만, 선구성의 아까시나무 유목과 기존 개체의 게릴라번식, 그리고 매몰 종자에 의한 발아 등으로 아까시나무림은 지속적으로 반복해서 유지되어 갈 것으로 예측되었다.
가죽나무는 소태나무과의 하록성 다년생 목본식물로 중국이 분포 중심지인 식물로 알려져 있다. 가죽나무는 생장이 매우 빠르고 유성생식과 무성생식이 가능하다. 또한, 바람이나 물에 의해 모체로부터 멀리 떨어진 장소까지 산포가 가능하다(Kowarik and Saumel, 2008). 본 식물은 중금속으로 오염된 토양환경과 대기오염도가 높은 지역 등 식물체가 번성하기 힘든 가혹한 환경조건에 대한 뛰어난 내성을 나타낸다. 그리고, 다른 식물의 발아, 생장, 생식을 억제하는 광합성 이차 대사산물을 만들며, 외래곤충인 꽃매미의 주요 기주식물로 여러 가지 생태적인 문제점을 일으키는 것으로 보고되고 있다(Lee et al., 2009).
가죽나무-아까시나무-참오동나무군락 고정조사구 3의 층위별 상대우점치 분석결과 교목층을 형성하였던 가죽나무, 참오동나무는 모두 도태된 상태였다. 교목층에서는 아까시나무가 상대우점치 100.0%로 우점하였으며 아교목층에는 출현종이 없었다. 관목층에서는 구기자나무가 일부 생육하고 있었다. 종수 및 개체수 분석결과 2종 38개체로 매우 적었으며 이에 따라 샤논의 종다양도지수 또한 0.1894로 매우 낮았다(Table 8 참조). 식물군집구조 분석결과를 종합해보면 교목층은 아까시나무로 우점하고 있으며 아교목층, 관목층 모두 미발달된 상태로 장기간 아까시나무림으로 유지될 것으로 판단되었다.
가죽나무-아까시 나무-참오동나무군락의 식생 변화 경향을 분석한 결과 목본 종수는 2008년까지 7-11종을 유지하였으나 2012년 5.5종, 2016년 6종에서 2021년 2종으로 크게 감소하였다. 이러한 변화는 교목층 세력 도태로 인한 것으로 판단되며 이후 유입종들에 의해 종수는 지속적으로 변화할 것으로 판단되었다. 교목층과 아교목층 개체수의 경우 2008년까지 교목층 7.5-10.0개체, 아교목층 7.3-12.5개체가 유지되었으나 2012년 교목층 5.0개체, 아교목층 2.5개체, 2021년 교목층 3.0개체, 아교목층 3.0개체로 감소하였다(Table 9 참조). 샤논의 종다양도지수는 2008년까지 0.5477-0.7829로 유지되었으나 2012년부터 감소하기 시작하여 2021년 0.1894로 크게 감소하였다(Table 10 참조).
국내에는 37종의 버드나무류가 분류되어 있으며, 하변 연목림에는 하천유역, 입지 안정성, 수위 등에 따라 버드나무류가 우점하는 13개의 식물군락이 발달하고 있다(Lee, 2004). Kim and Lee(1998)는 버드나무류의 식물군락은 영양번식이 가능하고 척박한 토양에서도 가지와 뿌리의 생장이 빨라 신속히 토양을 안정화하는 특성이 있다고 하였다.
능수버들군락 고정조사구 4의 층위별 상대우점치 분석결과 아까시나무가 교목층에서 64.0%, 아교목층에서 80.8%로 우점하였고, 아까시나무 이외 수종으로 교목층에 능수버들이 23.2%, 버드나무가 12.8%, 아교목층에 층층나무가 19.2% 출현하였다. 관목층에서는 개나리가 40.2%, 찔레꽃이 29.8%로 높은 우점도를 보인 가운데 딱총나무, 아까시나무, 구기자나무, 담쟁이덩굴이 함께 출현하였다. 종수 및 개체수 분석결과 9종 108개체로 다양하였으며 이에 따라 샤논의 종다양도지수는 0.7591으로 능수버들군락에 설정된 다른 조사구에 비해 다소 높은 수치를 보였다(Table 11 참조). 식물군집구조 분석결과를 종합해보면 교목층과 아교목층은 아까시나무로 우점하고 있으며 관목층에 교목성상의 수종이 없어 버드나무와 능수버들이 도태되며 아까시나무림으로 유지될 것으로 판단되었다.
능수버들군락 고정조사구 5의 층위별 상대우점치 분석결과 교목층, 아교목층은 아까시나무 1종이 상대우점치 100.0%로 우점하였고 관목층에서는 찔레꽃이 62.6%로 높은 우점도를 보인 가운데 조릿대, 개나리, 딱총나무, 말발도리가 함께 출현하였다. 종수 및 개체수 분석결과 6종 145개체로 종수는 능수버들군락에 설정된 다른 조사구와 유사하였으나 개체수는 비교적 많았다. 샤논의 종다양도지수는 0.6263로 타 조사구와 유사하였다(Table 12 참조). 식물군집구조 분석결과를 종합해보면 교목층, 아교목층에서 아까시나무가 우점하고 있었으며 관목층에 교목성상의 수종이 없어 아까시나무림으로 유지될 것으로 판단되었다.
능수버들군락 고정조사구 6의 층위별 상대우점치 분석결과 교목층, 아교목층에서 아까시나무가 상대우점치 100.0%, 82.9%로 우점하였고 아까시나무 이외 수종으로 아교목층에 복사나무가 17.1% 출현하였다. 관목층에서는 찔레꽃이 54.6%로 높은 우점도를 보인 가운데 칡, 딱총나무, 까마귀밥나무, 아까시나무, 복사나무가 함께 출현하였다. 종수 및 개체수 분석결과 6종 106개체로 종수는 능수버들군락에 설정된 다른 조사구와 유사하였다. 샤논의 종다양도지수도 0.6462로 타 조사구와 유사하였다(Table 13 참조). 식물군집구조 분석결과를 종합해보면 교목층, 아교목층에서 아까시나무가 우점하고 있었으며 관목층에 교목성상의 수종이 없어 아까시나무림으로 유지될 것으로 판단되었다.
능수버들군락의 식생 변화 경향을 분석한 결과 목본 종수는 2008년까지 9-12종을 유지하였으나 2012년 3.6종으로 감소 이후 2016년에는 11종, 2021년 10종으로 다시 증가하였다. 이러한 변화는 2010년 태풍에 의한 풍도목 발생과 버드나무의 쇠퇴로 인한 것으로 판단되며 이후 교목층의 밀도 감소에 따라 유입종들에 의한 종수 변화는 지속될 것으로 판단되었다. 층위별 개체수의 경우 교목층과 아교목층은 2016년까지 지속적으로 감소하다 2021년 증가하는 경향을 보였다. 관목층은 이러한 교목층과 아교목층의 개체수 변화에 따라 상층이 개방되면서 개체수가 증가하였다(Table 14 참조). 샤논의 종다양도지수는 종수가 급감한 2012년까지 0.4205로 크게 감소하였으나 2016년 다시 0.8295로 예전 수준으로 회복되어 2021년 0.8205로 나타났다(Table 15 참조).
자연발생 외래목본인 아까시나무 세력이 전반적으로 증가하고, 자연발생 자생목본인 버드나무와 능수버들은 세력이 감소하였다. 아까시나무의 경우 태풍 피해로 인해 전체적인 세력이 감소하였으나 아교목층과 관목층에 지속적으로 발생하여 회복되고 있는 것으로 판단되었다. 버드나무류의 경우 지속적인 건조한 환경, 열악한 생육기반 등으로 인해 쇠퇴가 예측되었다. 사면지역의 자연발생숲은 이후 지속적으로 모니터링하여 매립지 사면의 식생천이 방향을 예측할 필요가 있으며, 숲의 안정화를 방해하는 요인인 덩굴식생의 관리와 유입되는 수종으로 인한 종구성 변화 등을 지속적으로 확인해야 할 것이다. 조경수식재지는 2012년 0.8%에 불과하였으나 아까시나무 도복지, 덩굴식물 분포지, 초본식생지역 등을 대상으로 지속적으로 확대하여 7.6%로 증가하였다. 조경수식재지는 상수리나무, 왕벚나무, 팥배나무, 꽃복숭아나무 등 다양한 수종을 식재하여 관리하고 있으며, 대체로 생육상태가 양호하였다. 유입종들에 의해 변화가 나타나고 있었으며 주변으로부터 유입되는 외래종 아까시나무, 가시박, 단풍잎돼지풀에 의한 위협이 있어 지속적인 관리와 모니터링이 필요하며, 향후 매립지 사면에 적합한 수종 개발, 식생복원기법을 제시하기 위한 기초자료 구축이 필요하다.
아까시나무군락은 2010년 이후 목본 종수 감소 후 다시 증가 추세를 보였다. 2012년 이후 관목이 성장하면서 교목과 아교목층 개체수는 증가하였고, 관목층은 개체수를 유지하는 것으로 분석되었다. 향후 아까시나무군락은 아까시나무림으로 유지될 것으로 예측되었다. Wei and Oh(2021) 연구에 따르면 노을공원 사면 아까시나무군락은 뽕나무가 경쟁종으로 유입이 된 후 상․중층에서 세력이 커지면서 아까시나무의 상대우점치가 감소하는 것으로 나타났고, 일부 지역에서는 교란이 발생되어 아까시나무와 능수버들의 경쟁이 발생하고 있다. 본 연구 결과와 다른 이유는 조사구 위치 차이에 따른 결과로 보이며, 현존식생 조사 결과 뽕나무의 세력은 지속적으로 확장되고 있음을 확인하였다. 가죽나무-아까시나무-참오동나무군락의 경우 목본 종수 및 개체수는 2008년까지 증가하다가 2012년부터 교목층, 아교목층, 관목층에서 모두 감소하였다. 현재 교목층 참오동나무와 가죽나무는 도태된 상태로 아까시나무림으로 유지될 것으로 예측되었다. 능수버들군락은 목본 종수가 2012년 감소하였다가 2016년부터 증가하였다. 아까시나무림으로 천이된 상태였으며 일부 조사구에서만 교목층에 아까시나무와 함께 능수버들이 생육하고 있으나 대부분 쇠퇴한 상태이었다. 향후 능수버들 쇠퇴 후 관목층의 아까시나무가 증가하는 것으로 보아 아까시나무림으로 천이가 예측되었다. 사면지역 주요 군락은 출입통제를 지속적으로 유지해야 하며 고사목의 발생과 같이 다른 생물의 유입을 유도할 수 있는 환경인자들은 제거하지 않고 유지해야 한다. 그리고 이 식생 군락은 월드컵공원 사면지구의 대표식생으로서 정밀 모니터링을 통해 군락별 생태계 변화를 파악하여야 한다. 이는 매립지 사면지구의 자연발생숲의 변화 예측 및 생태계 복원 차원의 중요한 기초자료로 활용될 수 있다.
4. 결론
본 연구는 월드컵공원 사면지구를 대상으로 23년간(1999-2021년)의 현존식생과 식물군집구조 조사를 통해 식생 현황 및 변화 특성을 분석하였다. 자연발생 외래목본인 아까시나무 세력이 전반적으로 증가하고, 자연발생 자생목본인 버드나무와 능수버들은 세력이 감소하였다. 아까시나무는 태풍 피해로 인해 전체적인 세력이 감소하였으나 아교목층과 관목층에 지속적으로 발생하여 회복되고 있는 것으로 판단되었다. 버드나무류의 경우 지속적인 건조한 환경, 열악한 생육기반 등으로 인해 쇠퇴가 확인되었다. 아까시나무-능수버들군락의 경우 능수버들이 모두 도태되고 아까시나무림으로 변화가 예측되었다. 사면지구는 아까시나무, 가시박, 단풍잎돼지풀에 의한 자생식생 교란 위협이 있어 지속적인 관리와 모니터링이 필요하며, 향후 매립지 사면에 적합한 수종 개발, 식생복원 기법의 기초자료 구축이 필요하다.
아까시나무군락은 2010년 이후 목본 종수 감소 후 다시 증가 추세를 보였다. 2012년 이후 관목이 성장하면서 교목과 아교목층 개체수는 증가하였고, 관목층은 개체수를 유지하는 것으로 분석되었다. 향후 아까시나무군락은 아까시나무림으로 유지될 것으로 예측되었다. 가죽나무-아까시나무-참오동나무군락의 경우 목본 종수 및 개체수는 2008년까지 증가하다가 2012년부터 교목층, 아교목층, 관목층에서 모두 감소하였다. 현재 교목층 참오동나무와 가죽나무는 도태된 상태로 아까시나무림으로 유지될 것으로 예측되었다. 능수버들군락은 목본 종수가 2012년 감소하였다가 2016년부터 증가하였다. 아까시나무림으로 천이된 상태였으며 일부 조사구에서만 교목층에 아까시나무와 함께 능수버들이 생육하고 있으나 대부분 쇠퇴한 상태이었다. 관목층의 아까시나무가 증가하는 것으로 보아 아까시나무림으로 천이가 예측되었다. 사면지역의 자연발생숲은 이후 지속적인 변화를 모니터링하여 향후 매립지 사면의 식생형성 방향을 예측할 필요가 있으며, 숲의 안정화를 방해하는 요인인 덩굴식생의 지속적인 관리와 유입되는 수종으로 인한 종 구성 변화 등을 지속적으로 확인해야 할 것이다.
본 연구의 한계로 월드컵공원 매립지 사면지역 토양과 수목 생육과의 상관관계를 밝힐 필요가 있었으며, 기온 및 강수량과 같은 기상요인에 따른 식생 분포 변화를 시계열적으로 분석하지 못하였다. 또한 식생 관리방안은 현황을 바탕으로 지역별 식생기반 및 식생구조 개선방안을 세부적으로 도출할 필요가 있다.