




I. 서론
산업화․도시화로 인한 자연훼손은 심각한 녹지 환경의 감소를 초래하였으나, 최근 식물이 인간에게 주는 다양한 효과로 인해 녹지공간을 확대하고자 노력하고 있다. 옥외공간은 수목원, 공원, 정원, 가로수 등의 형태로 녹지면적을 확대하고 있으며, 건축물에서는 옥상녹화, 벽면녹화, 실내정원 등으로 녹지면적을 확보하려고 노력하고 있다. 특히 현대인들은 하루의 80~90%에 해당하는 시간을 실내에서 보내고 있으며(Statistics Korea, 2009), 이에 따라 실내식물이 인간에게 미치는 영향에 대한 연구도 꾸준히 이루어지고 있다. 미국항공우주국(National Aeronautics and Space Administration)은 우주선이라는 극단적인 밀폐환경에서 인간의 생존과 쾌적한 생활에 대해 다방면으로 연구하였으며, 실내에 식물을 배치한 결과, 적정 실내 온도와 습도를 유지하고, 더불어 실내 오염물질을 효율적으로 제거한다고 보고하였다(Wolverton et al., 1989). 실내식물은 현대인들에게 심리적인 안정감을 주며, 스트레스를 경감하고(Son et al., 1997), 창조적인 작업에 긍정적인 영향을 주는 것으로 알려졌다(Shibata and Suzuki, 2002). 실내에 녹지공간 조성을 위한 노력이 지속적으로 이루어지고 있으나, 실내 환경 특성상 식물 생존에 절대적인 광환경은 기존의 실내조명만으로는 한계가 있고(Son, 2013), 이용 식물별로 생육에 알맞은 적절한 광량이 다르기 때문에, 실내조경에서는 실내 광조건에 적합한 식물을 선택하거나, 생육적 특성을 고려한 광량 조절이 필요하게 된다(Choi and Lee, 2005; Song et al., 2013). 실내 조경식물로 사용되는 대부분 관엽식물들은 열대 및 아열대 지방의 나무 그늘 아래에서 자라는 내음성이 강한 식물류가 80% 이상을 차지하고 있다(Chen et al., 2003; Son, 2013). 이러한 관엽식물들은 주로 차광처리된 온실에서 재배되어 실내 환경 에 적응하는 일련의 순화과정을 거치게 된다(Chen et al.., 2003). Lee and Suh (1999)는 실내조경 식물소재 개발을 위해 Ardisia속 식물을 이용하여 자연광과 차광처리를 실시한 후, 식물생육을 연구하였고, Bang et al.(2000)은 백열등, 수은등 등 5종 광원에 따른 실내지피식물의 생육 반응에 대해 조사하였다. Park et al.(2010)은 실내환경 개선에 적합한 식물 선발을 위한 관엽식물들의 광-광합성반응을 연구하였으며, Heo et al. (2010)은 발광다이오드(Light Emitting Diode) 청색, 적색, 단일광, 혼합광을 이용하여 디펜바키아와 피커스의 온실재배 시 보광광원으로 적합 여부에 대해서 연구하였다. 기존의 연구들은 자연광 차광 및 실내식물 재배를 위한 광조건에 대한 연구가 다수였으며, 낮은 실내 광환경에서의 식물 유지 관리를 위한 생육에 대한 연구는 아직 미비한 실정이다. 또한, 다양한 식물을 실내조경에 이용하고자 관엽식물 위주의 식물선정에서 벗어나 이끼, 양치식물, 목본류(Ju et al., 1999; Lee and Lee, 2013; Kwon and Park, 2017) 등의 식물에 대한 연구도 진행되고 있다. 그러나 이끼, 양치식물 등은 겨울철 실내 난방으로 인한 건조한 환경에서 면밀한 관리가 필요하며, 아교목, 교목 등 키가 큰 수종들은 일반적으로 실내공간에서는 공간적인 제약을 받을 수밖에 없다. 따라서 상록성 관목, 내음성이 강한 수종, 천천히 자라는 수종, 공기정화 효과 등 친환경적 기능성이 높은 수종 등의 한정된 범위에서 목본식물 선택이 필요하다. 이에 본 연구는 산호수, 팔손이와 크루시아 등 목본식물 3종을 대상으로 광량에 따른 생육반응을 구명하고자 수행하였다.
II. 재료 및 방법
실내녹화에 많이 사용되는 산호수(Ardisia pusilla), 크루시아(Clusia rosea), 팔손이(Fatsia japonica)를 식물재료로 이용하였다. 산호수는 자금우과(Myrsinaceae)에 속하는 상록성, 포복형 식물로 번식이 용이하고, 비교적 따뜻한 기후에서 잘 자라며, 실내 베란다에서 월동이 가능하다. 팔손이는 두릅나무과(Araliaceae)과에 속하는 국내에 자생하는 상록관목성 식물로 손쉽게 구할 수 있는 장점이 있으며, 공기정화능력이 좋은 식물로 알려져 있다(Song, 2012). 크루시아는 물레나무과(Guttiferae) 계통으로 북미에서 멕시코, 카리브지역에 광범위하게 분포하며, 약 200여종이 있다(Wagner et al., 1999). 상록 활엽 소관목으로 C3와 CAM으로 광합성을 하며, 관수관리가 용이하다(Schmitt et al., 1988; Kwon and Park, 2017). 청주 근처 농원(양촌화훼, 남이면)에서 초고가 유사한 3치 포트 식물을 일괄 구입 후, 원예용 상토(Wonjo Mix, Nongkyung, Korea)로 분갈이한 다음 실내온도 약 25℃로 조절된 실험실내에서 순화하였다. 실내식물의 녹피율이 높을수록 심리적 안정감, 공기 정화 등에 효과적이다. 한정된 실내공간에서 녹피율을 높이는 방법으로 벽면녹화가 많이 사용되며, 벽면녹화에는 작은 치수의 포트 식물이 일반적으로 식재되고 있다. 관수는 식물이 수분스트레스를 받지 않을 정도로 주 1회 저면관수하였다. 실험은 2014년 5월부터 9월까지 총 120일간 진행하였다.
일반적으로 우리나라에서는 밝기의 정도를 룩스(Lux, lx)의 단위로 표현한다. 식물의 경우는 빛에 포함된 광량자 하나를 받아들여 하나의 광합성을 하기 때문에 광양자속밀도(Photosynthetic Photon Flux Density: PPFD)라는 표현을 쓴다. 이는 단위시간과 단위면적당 입사된 광량자수를 의미하며, 이때의 빛의 강도를 나타내는 방법이다. 노지에서 재배하는 토마토의 광보상점과 광포화점은 각각 3,000lx, 70,000lx이며, 광량자속밀도는 각각 36µmol․m−2․s−1, 847µmol․m−2․s−1로 환산된다. 차광막을 씌워 재배하는 인삼의 광보상점과 광포화점은 각각 500lx, 12,000lx 이며, 광량자속밀도는 각각 6µmol․m−2․s−1, 145µmol․m−2․s−1로 환산된다(Lee and Kim, 2012). 8종의 실내 관엽식물들의 광합성 반응을 알아보고자 수행한 선행연구에서는 식물들이 광포화점이 100µmol․m−2․s−1 전후에서 나타났다고 보고하여(Park et al., 2010), 이를 토대로 본 실험에서는 광합성 광량자속밀도를 15, 30, 60, 120µmol․m−2․s−1의 4수준으로 설정하였다. 90% 차광막으로 외부광 유입을 차단하고, 형광등을 이용해 보광한 다음, 식물 정단부로부터 광량측정기(Li-1400, LI-COR, USA)로 4수준의 광량을 조절하였다. 광조사 시간은 12/12시간(명기/암기)으로 자동조절하였다.
광량에 따른 생육상태를 파악하기 위해, 실험기간 동안 4주 간격으로 초고, 엽장, 엽폭, 엽록소량, 엽록소형광 등의 생육측정을 실시하였다. 초고는 식물의 정단부까지의 길이를 측정하였다. 엽장, 엽폭은 정단부 아래 5cm 이내의 가장 크고 신선한 잎을 선정하여 측정하였다. 엽면적, 생체중, 건조중은 실험 종료 후 측정하였다. 엽면적은 엽면적 측정기(Li-3000A, LI-COR, USA)를 이용하여 모든 잎을 측정하여 합산했다. 건조중은 열풍순환식 건조기(HB-502M, Hanbaek science, Korea)를 이용하여 70℃의 온도에서 72시간 동안 건조시킨 후 측정하였다. 엽록소함량은 엽록소측정기(SPAD-502Plus, Konica Minolta, Japan)를 이용하여, 엽색은 색차계(CR-300, Minolta, Japan)를 이용하여 잎의 앞면에서 엽맥을 피해 Hunter 값(L, a, b)을 측정하였다. 환경스트레스에 대한 내성을 파악하기 위해, 엽록소형광측정기(PAM-2000, Walz, Germany)를 사용하여 엽록소형광을 개체 당 3회 측정하였다. 엽록소함량, Hunter값과 엽록소형광은 엽폭과 엽장을 측정한 잎에서 측정하였다.
III. 결과 및 고찰
실험 종료 후 광량별 식물의 생육 상태에서, 3종 식물 모두 총엽면적은 120µmol․m−2․s−1에서 가장 넓었다. 특히, 산호수는 줄기 생장이 컸으며, 크루시아는 잎이 가장 넓었다(Figure 1 참조).

초고 변화는 3종 식물 모두 120µmol․m−2․s−1에서 가장 큰 것으로 나타났다. 산호수는 120µmol․m−2․s−1에서 실험종료 후 평균 초고 차이가 10.8cm로 가장 컸으며, 나머지 광량에서는 1.7~2.5 cm의 차이를 보였다. 크루시아는 60과 120µmol․m−2․s−1에서 크게 성장하는 경향을 보였으나, 15와 30µmol․m−2․s−1에서는 거의 생장하지 않는 것으로 나타났다. 실험종료 후 평균 초고 변화량은 15, 30, 60, 120µmol․m−2․s−1에서 각각 1.5, 2.8, 12.5, 16.9cm로 나타났다. 팔손이는 15, 30, 60, 120µmol․m−2․s−1에서 각각 2.5, 5.6, 8.9, 10.8cm로 나타나, 광량이 높을수록 초고 성장폭이 큰 것으로 나타났다(Figure 2 참조).
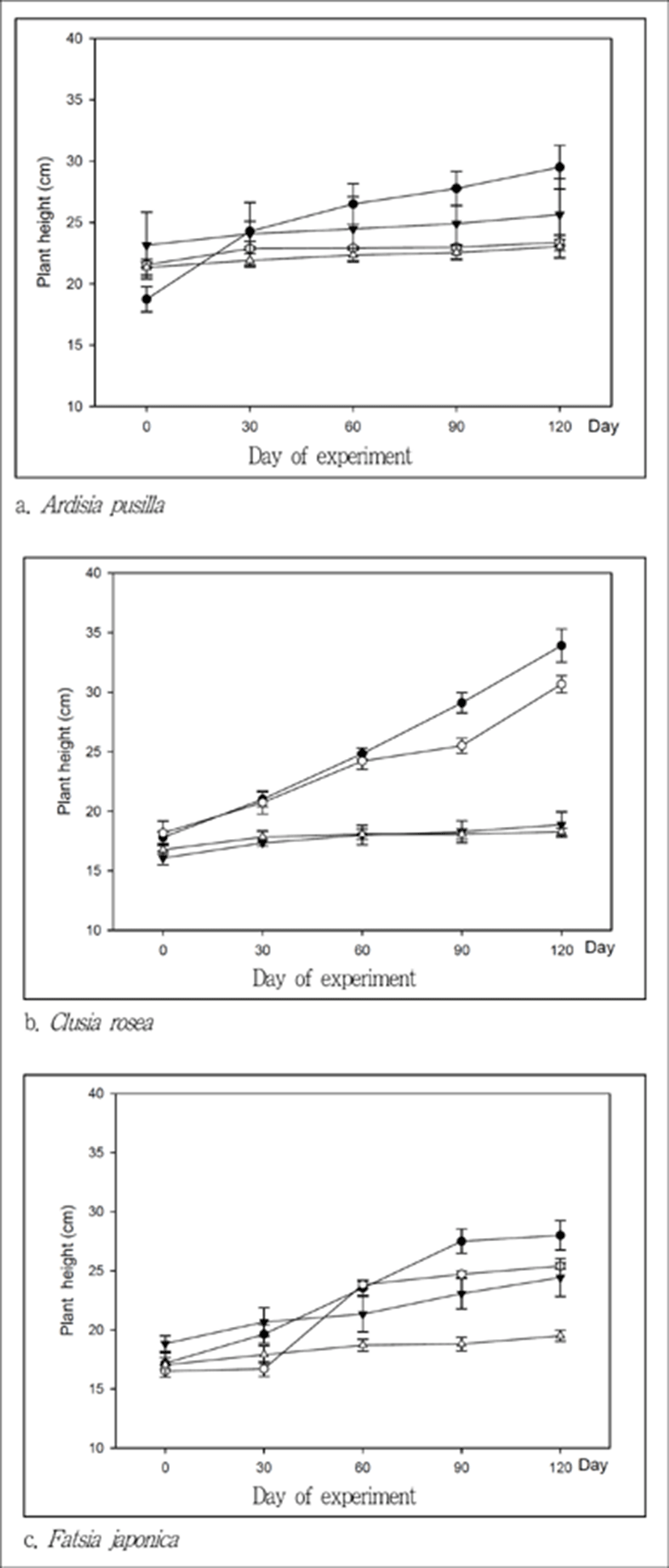
산호수의 초고, 엽장, 엽폭은 광량 차이에 따른 유의적인 차이가 없었으며, 총엽면적은 광량이 높을수록 높아지는 경향을 보였다(Table 1 참조). 이는 자금우를 이용한 차광실험에서도 엽장과 엽폭은 광량에 따른 유의적인 차이가 없었으나, 엽면적은 차광 35%, 차광 75%, 자연광 수준으로 높았던 것과 유사한 결과가 나타나(Lee and Suh, 1999), 자금우과 식물인 자금우, 산호수는 광량이 엽수에 영향을 미치는 것으로 판단된다. 크루시아 초고는 60, 120µmol․m−2․s−1과 15, 30µmol․m−2․s−1에서 유의적인 차이를 보였다. 엽장과 엽폭은 산호수와 마찬가지로 광량에 따른 유의적인 차이가 없었으나, 총엽면적은 광량이 높을수록 높게 나타났다. 크루시아는 광질에 따른 초고는 유의적인 차이가 있었으나, 엽장, 엽폭은 유의적인 차이가 나타나지 않았다고 보고하고 있어(Kim et al., 2013), 이 식물은 광질이나 광량이 측지 발달과 엽수 증가에 영향을 미치는 것으로 판단된다. 팔손이의 초고는 15, 30, 60µmol․m−2․s−1와 120µmol․m−2․s−1 사이에서는 유의적인 차이가 나타났다. 엽장, 엽폭, 엽면적은 광량에 따른 유의적인 차이가 나타났으며, 광량이 높아질수록 높게 나타나, 광량에 영향을 받는 것으로 판단된다.
생체중 및 건조중은 3종 식물 모두 광량에 따른 유의적인 차이를 보였고, 120µmol․m−2․s−1에서 지상부, 지하부의 생체중과 건조중 모두 높게 나타났다(Table 2 참조). 산호수는 광량에 따른 엽면적의 차이가 생체중 및 건조중의 유의적인 차이로 이어졌으며, 지하부도 생체중과 건조중에서 유의적인 차이를 보였다. 크루시아는 초고와 엽면적이 지상부의 생체중과 건조중에 모두 반영되어, 120, 60µmol․m−2․s−1 순으로 높게 나타났으며, 15, 30µmol․m−2․s−1에서는 유의적인 차이가 나타나지 않았다. 크루시아는 120µmol․m−2․s−1에서 지하부의 생체중과 건조중이 가장 높게 나타났다. 팔손이의 지상부 생체중은 120, 60, 30, 15µmol․m−2․s−1 순으로 높았고, 지상부 건조중, 지하부 생체중과 건조중은 15와 30µmol․m−2․s−1에서는 유의적인 차이가 없었으나, 60µmol․m−2․s−1 이상에서는 유의적인 차이가 나타났다.
식물은 고온과 저온, 강광, 약광, 건조, 침수 등의 환경에 대한 스트레스를 받는다. 일시적이거나 가벼울 경우는 회복이 가능하며, 또한 내성이 생기기도 한다. 환경 스트레스는 직․간접적으로 광계 II (Photosystem II)와 같은 광합성 기구에 손상을 줄 수 있다. 광계 II의 반응을 조사하기 위해서는 엽록소형광 분석이 다양하게 이용되고 있으며, 이는 환경 스트레스에 대한 식물의 생리적 반응 및 광합성 기구에 대한 다양한 정보를 제공한다(Oh et al., 2014). 최대 양자수율(Fv/Fm)은 광계Ⅱ 반응센터가 닫힌 상태의 최대형광(Maximal Fluorescence, Fm)과 광계Ⅱ 반응센터가 열린 상태의 최소형광(Minimal Fluorescence, Fo)을 측정하여 Fv=Fm-Fo의 식으로 산출한다(Van Kooten and Snell, 1990). 엽록소형광 Fv/Fm은 식물체의 스트레스를 비파과적으로 측정이 가능하며, 일반적으로 0.78~0.84범위에 있으면 스트레스가 없는 것으로 간주한다(Yoo et al., 2012). 산호수는 광량이 15, 30µmol․m−2․s−1로 비교적 낮을 때 Fv/Fm값이 0.786으로 나타나, 정상범위에 있었으나, 60, 120µmol․m−2․s−1에서는 정상 범위를 벗어나고 있어 약간의 스트레스를 받고 있는 것으로 나타났다(Table 3 참조). 실내조경 식물소재 개발을 위한 자생 Ardisia속의 식물 연구(Lee and Suh, 1999)에서 음지식물인 산호수의 7월초 자생지의 광량은 27.8~92.6µmol․m−2․s−1로 나타나, 산호수가 스트레스를 받지 않고 생육할 수 있는 광량은 60µmol ․m−2․s−1미만일 것으로 판단된다. 크루시아와 팔손이는 모든 광량에서 Fv/Fm값이 정상 범위에 있어, 산호수보다는 광량에 대한 민감도가 낮은 경향을 보였다. 인삼의 광포화점은 145µmol․m−2․s−1으로(Lee and Kim, 2012), 본 실험에서 광량의 범위가 15~120µmol․m−2․s−1이므로, 가장 높은 수준인 120µmol․m−2․s−1는 인삼밭 차광막 아래의 환경과 비슷할 것으로 생각된다. 크루시아, 팔손이는 광량 4수준의 범위에서 Fv/Fm이 모두 정상범위로 실내광량에 따른 실내식물 선정 시 선택의 폭을 넓힐 수 있을 것으로 판단된다.
SPAD값은 산호수와 팔손이는 광량에 따른 유의적인 차이가 없었으나, 크루시아는 광량에 따른 유의적인 차이를 보였으며, 특히, 120µmol․m−2․s−1에서 가장 낮은 값을 나타냈다. 식물은 낮은 광량에서는 광합성의 효율을 높이기 위해 엽록소 함량이 증가하는 것으로 알려져 있다(Lee et al., 2013). 120 µmol․m−2․s−1에서 크루시아는 반대의 경우로, 초장의 성장폭이 가장 높아 실험 종료시 식물의 정단부가 광원에 근접하여 SPAD값이 유의적으로 낮게 나타난 것으로 판단된다. Jeong and Kim (1999)의 목본식물인 송악과 수호초의 차광처리 연구에서는 차광량이 높아질수록 엽록소 a, b의 함량이 다소 높다고 하였으나, 본 연구에서 측정한 산호수, 팔손이는 SPAD값은 광량에 따른 차이가 없었던 것은 Jeong and Kim (1999)의 직사광선 차광보다도 낮은 광량 범위에서 실험되었기 때문으로 판단된다.
엽색지수를 위한 색차계 측정값은 엽색의 밝기(L: 0~100), 녹색-적색 기울기(a: -100~+100), 청색-황색기울기(b: -100 ~+100)를 나타낸다. 산호수는 광량에 따른 L, a, b 값의 유의적인 차이가 없었다. 크루시아의 경우, 15, 30µmol․m−2․s−1에서는 밝기와 청황색의 변화는 유의적인 차이가 없었으나, 60, 120mol․m−2․s−1에서는 유의적인 차이가 나타났다. 녹적색 기울기는 60µmol․m−2․s−1에서 가장 녹색이 많이 발현되었으며, 그 다음으로 120, 30, 15µmol․m−2․s−1 순으로 높게 나타났다. 팔손이는 120µmol․m−2․s−1에서 가장 밝은 색을 나타냈으며, 녹적색 기울기는 30, 15, 60, 120µmol․m−2․s−1 순으로 높았고, 청황색 기울기는 120, 60, 15, 30µmol․m−2․s−1순으로 높았다. 일반적으로 엽록소 함량은 저광량 조건에서 높게 나타나고, 엽록소 함량이 많아지면 엽색이 짙어지는 경향을 보이는데(Simpson and Lee, 1976), 본 연구에서도 크루시아는 광량이 가장 높은 120µmol․m−2․s−1에서 가장 낮은 SPAD값을 나타내었고, 15, 30µmol․m−2․s−1에서 L값이 낮아 짙은 엽색을 나타내었다. 팔손이는 SPAD값은 유의적인 차이를 나타내지 않았으나, 크루시아와 마찬가지로 15, 30µmol ․m−2․s−1에서 L값이 낮아 짙은 엽색을 나타내었다. 15µmol ․m−2․s−1에서 3종 식물의 L값은 25.5~27의 범위에 있었으나, 광량이 높아질수록 L값의 차이가 커져서 120µmol․m−2․ s−1에서 L값은 산호수 25.5, 크루시아 32.3, 팔손이 43으로 17.5의 차이를 보였다. a, b값에서도 광량이 높을수록 식물간의 차이가 커졌다. 이는 일정한 광도로 설정된 실내공간에서, 실내 식물의 혼합식재 시 식물형태뿐만 아니라, 광량에 따른 식물의 명암, 적색도, 황색도의 차이로 단조로운 녹색에서 그러데이션의 변화가 가능할 것으로 판단되며, 넓은 벽면 녹화 등에서 다양한 작품표현에 도움이 될 것으로 기대된다.
IV. 결론
실내식물 도입의 다양성을 위해 목본식물인 산호수, 크루시아, 팔손이 3종을 선정하여 15, 30, 60, 120µmol․m−2․s−1 의 4수준의 광량에서 120일간 초고, 엽장, 엽폭, 엽면적, 지상부와 지하부의 생체중 및 건조중, 엽록소형광(Fv/Fm), SPAD, 엽색을 나타내는 Hunter 값 L, a, b를 측정하였다.
산호수는 13개의 측정항목 중 Fv/Fm, 지상부 생체중, 건조중, 지하부 건조중에서 유의적인 차이를 보였다. 120µmol․m−2․s−1에서 엽면적, 지상부와 지하부의 생체중 및 건조중이 가장 높은 것으로 나타났다. Fv/Fm 값은 60, 120 µmol․m−2․s−1에서 정상범위를 벗어나, 광량에 대한 스트레스를 받고 있는 것으로 나타났다. 크루시아는 13개의 측정항목 중 엽장, 엽폭, Fv/Fm를 제외한 모든 항목에서 유의적인 차이가 나타났다. 광량이 높아질수록 초고, 엽면적, 지상부와 지하부의 생체중 및 건조중에서 높은 값이 나타났다. 일반적으로 성장곡선은 일정 시간 후 완만해지는 경향을 나타내는데, 크루시아는 실험기간 동안 60, 120µmol․m−2․s−1에서는 지속적인 성장을 하여, 추후 적정 광량을 찾기 위한 추가적인 실험이 수행되어져야 할 것으로 판단된다. 120µmol․m−2․s−1에서 가장 낮은 SPAD값을 나타냈고, Hunter값인 L, a, b는 각각 60, 15, 60µmol․m−2․s−1에서 가장 높은 것으로 나타났다. 팔손이는 13개의 측정항목 중 엽폭, Fv/Fm, SPAD값을 제외한 모든 항목에서 유의적인 차이가 나타났다. 120µmol․m−2 ․s−1에서 가장 높은 초고 생장율을 보였으며, 광량이 높아질수록 초고, 엽장, 엽면적, 지상부와 지하부의 생체중 및 건조중이 높아지는 경향을 보였다. Hunter값 L, a, b는 각각 120, 30, 120µmol․m−2․s−1에서 가장 높은 것으로 나타났다.
한정된 실내공간에서 설치된 조경식물에서 특정식물의 지나친 초고, 엽장, 엽폭, 엽면적의 성장은 초기 조성된 조경작품의 형태를 심하게 변형시키며, 다른 식물의 광환경을 저해하는 요소가 될 수 있다. 크루시아는 60, 120µmol․m−2․s−1에서 지속적인 성장을 보였으며, 팔손이는 모든 광량에서 일정 기간이 지나면 초고 성장속도가 완만해지고, 광량이 높아질수록 엽장, 엽폭, 엽면적이 증대하는 것으로 나타났다. 특정 공간의 광환경과 이에 맞는 식물의 성장 특성도 고려하여 식물을 선정한다면 관리비용을 저감할 수 있을 것으로 판단된다. 또한, 광량에 따른 Fv/Fm값을 고려한다면 식물 스트레스로 고사한 식물의 교체 비용도 절감할 수 있을 것으로 기대된다.
따라서 본 연구를 통하여 실내공간에서 단독 식재의 단조로움을 해결하기 위한 혼합식재시 동일한 광환경에서 성장 속도가 비슷한 식물을 식재하여 전체의 조화로움을 계속 유지하고, 광량에 따른 식물의 엽색을 활용하여 다양한 색상을 표현할 수 있을 것으로 판단된다. 그러나, 더 많은 식물종을 대상으로 적정 광량을 구명하는 추가적인 실험과 더불어 장기적인 모니터링이 이루어져야 할 것이다.